Q1. 什麼是基因?
自1900年孟德爾定律重新發現後,「基因怎樣控制性狀」的問題引起了許多遺傳學家的濃厚興趣。經過他們的努力,又出現了一批重要成果。
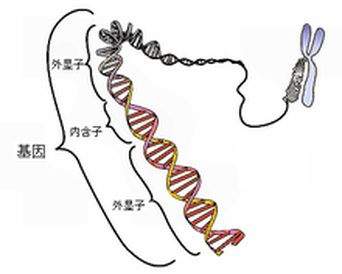
基因一詞來自希臘語,意思為「生」。是指攜帶有遺傳信息的DNA序列,是控制性狀的基本遺傳單位亦即一段具有功能性的DNA序列。基因通過指導蛋白質的合成來表達自己所攜帶的遺傳信息,從而控制生物個體的性狀表現。人類約有兩萬至兩萬五千個基因。
染色體在體細胞中是成對存在的,每條染色體上都帶有一定數量的基因。一個基因在細胞有絲分裂時有兩個對列的位點,稱為等位基因,分別來自父親與母親。按照其控制的性狀,又可分為顯性基因和隱性基因。
一般來說,生物體中的每個細胞都含有相同的基因,但並不是每個細胞中的每個基因所攜帶的遺傳信息都會被表達出來。不同部位和功能的細胞,能將遺傳信息表達出來的基因也不同。
人類基因組計劃 (human genome project, HGP) 是一項規模宏大,跨國跨學科的科學探索工程。其宗旨在於測定組成人類染色體 (指單倍體) 中所包含的30億個鹼基對組成的核苷酸序列,從而繪製人類基因組圖譜,並且辨識其載有的基因及其序列,達到破譯人類遺傳信息的最終目的。該計劃起始於1990年,已基本確定了人類的所有基因。
基因一詞來自希臘語,意思為「生」。是指攜帶有遺傳信息的DNA序列,是控制性狀的基本遺傳單位亦即一段具有功能性的DNA序列。基因通過指導蛋白質的合成來表達自己所攜帶的遺傳信息,從而控制生物個體的性狀表現。人類約有兩萬至兩萬五千個基因。
染色體在體細胞中是成對存在的,每條染色體上都帶有一定數量的基因。一個基因在細胞有絲分裂時有兩個對列的位點,稱為等位基因,分別來自父親與母親。按照其控制的性狀,又可分為顯性基因和隱性基因。
一般來說,生物體中的每個細胞都含有相同的基因,但並不是每個細胞中的每個基因所攜帶的遺傳信息都會被表達出來。不同部位和功能的細胞,能將遺傳信息表達出來的基因也不同。
人類基因組計劃 (human genome project, HGP) 是一項規模宏大,跨國跨學科的科學探索工程。其宗旨在於測定組成人類染色體 (指單倍體) 中所包含的30億個鹼基對組成的核苷酸序列,從而繪製人類基因組圖譜,並且辨識其載有的基因及其序列,達到破譯人類遺傳信息的最終目的。該計劃起始於1990年,已基本確定了人類的所有基因。
★基因的研究歷史★
美國實驗胚胎學家、遺傳學家湯馬士·亨特·摩根和他的學生們於1908年前後開始利用果蠅作了大量的潛心研究。他在1910年通過果蠅眼色突變性狀的遺傳實驗發現了伴性遺傳現象,第一次揭示出一種或多種遺傳特性與某一特定染色體的明確聯繫;他和他的同事們進一步通過大量的果蠅雜交實驗又發現了遺傳學的第三個基本規律——連鎖互換規律,從而繼承和發展了孟德爾的遺傳學說。他們為遺傳染色體學說最終提供了更充分、直接、可靠的證據,並認為染色體是盂德爾式遺傳性狀傳遞機理的物質基礎。1926年他的巨著《基因論》出版,從而建立了著名的基因學說,他還繪製了著名的果蠅基因位置圖,首次完成了當時最新的基因概念的描述,即基因以直線形式排列,它決定著一個特定的性狀,而且能發生突變並隨著染色體同源節段的互換而交換,它不僅是決定性狀的功能單位,而且是一個突變單位和交換單位。
摩根等人還認為,基因是遺傳的功能單位,它能產生特定的表型效應;基因又是一個獨立的結構單位。在同源染色體之間可以發生基因的互換,但交換隻能發生在基因之間而不是發生在基因之內;基因可以發生突變,由一個等位形式變為另一等位形式,因而基因又是突變單位。這就是20世紀40年代以前流行的所謂「功能、交換、突變」三位一體的基因概念。這種認識把基因與染色體聯繫起來,說明了基因的物質性,基因存在的場所及排列方式,基因從此就不再是一個抽象的概念了。當然這時人們仍然不了解基因的化學本質以及基因是如何控制生物性狀的。
從20世紀40年代起,人們開始注意基因與性狀的關係,即開始研究基因如何控制性狀的問題,1941年,比德爾和塔特姆以紅色鏈抱霉為材料進行生化遺傳研究。他們通過誘變獲得了多種胺基酸和維生素的大量營養缺陷突變體。這些突變基因不能產生某種酶,或只產生有缺陷的酶。例如,有一個突變體不能合成色氨酸是由於它不能產生色氨酸合成酶。於是,研究者提出了「一個基因一種酶」的假說,認為基因對性狀的控制是通過基因控制酶的合成來實現的。這一假說在20世紀50年代得到充分驗證,後來發現有些蛋白質不只由一種肽鏈組成,如血紅蛋白和胰島素,不同肽鏈由不同基因編碼,因而1941年比德爾和塔特姆提出一個基因一個酶學說,證明基因通過它所控制的酶決定著代謝中生化反應步驟,進而決定生物性狀。又提出了「一個基因一條多肽鏈」的假設。「一個基因一種酶」和「一個基因一條多肽鏈」理論的提出,大大促進了分子遺傳學的發展,人們急切期望能搞清楚基因的化學結構。1949年鮑林與合作者在研究鐮刀型細胞貧血症時推論基因決定著多肽鏈的胺基酸順序,這樣20世紀40年代末至20世紀50年代初,基因是通過控制合成特定蛋白質以控制代謝決定性狀原理變得清晰起來。
雖然DNA在細胞核中很早就被發現,但證明其為遺傳物質的決定性實驗是1944年艾弗里的肺炎雙球菌的轉化實驗。他和麥卡蒂等人發表了關於「轉化因子」的重要論文,首次用實驗明確證實:DNA是遺傳信息的載體。1952年赫希和蔡斯進一步證明遺傳物質是DNA而不是蛋白質。
這一實驗不僅證明了DNA是遺傳物質,揭示了遺傳物質的化學本質,也大大推動了對核酸的研究。1953年,美國分子生物學家詹姆斯·沃森和英國物理學家佛朗西斯·克里克根據威爾金斯和富蘭克林所進行的X射線衍射分析,提出了著名的DNA雙螺旋結構模型,進一步說明基因成分就是DNA,它控制著蛋白質合成。進一步的研究證明,基因就是DNA分子的一個區段。每個基因由成百上千個脫氧核苷酸組成,一個DNA分子可以包含幾個乃至幾千個基因。基因的化學本質和分子結構的確定具有劃時代的意義,它為基因的複製、轉錄、表達和調控等方面的研究奠定了基礎,開創了分子遺傳學的新紀元。
從20世紀40年代起,人們開始注意基因與性狀的關係,即開始研究基因如何控制性狀的問題,1941年,比德爾和塔特姆以紅色鏈抱霉為材料進行生化遺傳研究。他們通過誘變獲得了多種胺基酸和維生素的大量營養缺陷突變體。這些突變基因不能產生某種酶,或只產生有缺陷的酶。例如,有一個突變體不能合成色氨酸是由於它不能產生色氨酸合成酶。於是,研究者提出了「一個基因一種酶」的假說,認為基因對性狀的控制是通過基因控制酶的合成來實現的。這一假說在20世紀50年代得到充分驗證,後來發現有些蛋白質不只由一種肽鏈組成,如血紅蛋白和胰島素,不同肽鏈由不同基因編碼,因而1941年比德爾和塔特姆提出一個基因一個酶學說,證明基因通過它所控制的酶決定著代謝中生化反應步驟,進而決定生物性狀。又提出了「一個基因一條多肽鏈」的假設。「一個基因一種酶」和「一個基因一條多肽鏈」理論的提出,大大促進了分子遺傳學的發展,人們急切期望能搞清楚基因的化學結構。1949年鮑林與合作者在研究鐮刀型細胞貧血症時推論基因決定著多肽鏈的胺基酸順序,這樣20世紀40年代末至20世紀50年代初,基因是通過控制合成特定蛋白質以控制代謝決定性狀原理變得清晰起來。
雖然DNA在細胞核中很早就被發現,但證明其為遺傳物質的決定性實驗是1944年艾弗里的肺炎雙球菌的轉化實驗。他和麥卡蒂等人發表了關於「轉化因子」的重要論文,首次用實驗明確證實:DNA是遺傳信息的載體。1952年赫希和蔡斯進一步證明遺傳物質是DNA而不是蛋白質。
這一實驗不僅證明了DNA是遺傳物質,揭示了遺傳物質的化學本質,也大大推動了對核酸的研究。1953年,美國分子生物學家詹姆斯·沃森和英國物理學家佛朗西斯·克里克根據威爾金斯和富蘭克林所進行的X射線衍射分析,提出了著名的DNA雙螺旋結構模型,進一步說明基因成分就是DNA,它控制著蛋白質合成。進一步的研究證明,基因就是DNA分子的一個區段。每個基因由成百上千個脫氧核苷酸組成,一個DNA分子可以包含幾個乃至幾千個基因。基因的化學本質和分子結構的確定具有劃時代的意義,它為基因的複製、轉錄、表達和調控等方面的研究奠定了基礎,開創了分子遺傳學的新紀元。
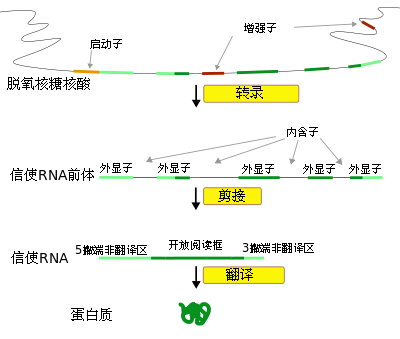
基因本質的確定為分子遺傳學發展拉開了序幕。1955年,美國分子生物學家本澤對大腸桿菌T4噬菌體作了深入研究,揭示了基因內部的精細結構,提出了基因的順反子(Cistron)概念。本澤把通過順反實驗而發現的遺傳的功能單位稱為順反子,1個順反子決定一條多肽鏈,順反子即是基因。1個順反子內存在著很多突變位點—--突變子,突變子就是改變後可以產生突變型表型的最小單位。1個順反子內部存在著很多重組子。重組子就是不能由重組分開的基本單位。理論上每一核苷酸對的改變,就可導致一個突變的產生,每兩個核苷酸對之間都可發生交換。這樣看來,一個基因有多少核苷酸對就有多少突變子,就有多少重組子,突變子就等於重組子。這個學說打破了過去關於基因是突變、重組、決定遺傳性狀的「三位一體」概念及基因是最小的不可分割的遺傳單位的觀點,從而認為基因為DNA分子上一段核苷酸順序,負責著遺傳信息傳遞,一個基因內部仍可劃分若干個起作用的小單位,即可區分成順反子、突變子和重組子。一個作用子通常決定一種多肽鏈合成,一個基因包含一個或幾個作用子。突變子指基因內突變的最小單位,而重組子為最小的重組合單位,只包含一對核苷酸。所有這些均是基因概念的偉大突破。 關於基因的本質確定後,人們又把研究視線轉移到基因傳遞遺傳信息的過程上。在20世紀50年代初人們已懂得基因與蛋白質間似乎存在著相應的聯繫,但基因中信息怎樣傳遞到蛋白質上這一基因功能的關鍵課題在20世紀60年代至20世紀70年代才得以解決。從1961年開始,尼倫伯格和科拉納等人逐步搞清了基因以核苷酸三聯體為一組編碼胺基酸,並在1967年破譯了全部64個遺傳密碼,這樣把核酸密碼和蛋白質合成聯繫起來。然後,沃森和克里克等人提出的「中心法則」更加明確地揭示了生命活動的基本過程。1970年霍華德·馬丁·特明以在勞斯肉瘤病毒內發現逆轉錄酶這一成就進一步發展和完善了「中心法則」,至此,遺傳信息傳遞的過程已較清晰地展示在人們的眼前。過去人們對基因的功能理解是單一的即作為蛋白質合成的模板。但是1961年法國弗朗索瓦·雅各布和雅克·莫諾的研究成果,又大大擴大了人們關於基因功能的視野。他們在研究大腸桿菌乳糖代謝的調節機制中發現了有些基因不起合成蛋白質模板作用,只起調節或操縱作用,提出了操縱子學說。從此根據基因功能把基因分為結構基因、調節基因和操縱基因。
★基因的種類★
★結構基因和調控基因:
根據操縱子學說,並不是所有的基因都能為肽鏈進行編碼。於是便把能為多肽鏈編碼的基因稱為結構基因,包括編碼結構蛋白和酶蛋白的基因,也包括編碼阻遏蛋白或激活蛋白的調節基因。有些基因只能轉錄而不能轉譯,如tRNA基因和rRNA基因。還有些DNA區段,其本身並不進行轉錄,但對其鄰近的結構基因的轉錄起控制作用,被稱為啟動基因和操縱基因。啟動基因、操縱基因與其控制下的一系列結構基因組成一個功能單位叫做操縱子(operon)。就其功能而言,調節基因、操縱基因和啟動基因都屬於調控基因。這些基因的發現,大大拓寬了人們對基因功能及相互關係的認識。
★斷裂基因:
20世紀70年代中期,法國生物化學家查姆幫(Chamobon)和波蓋特(Berget)在研究雞卵清蛋白基因的表達中發現,細胞內的結構基因並非全部由編碼序列組成,而是在編碼序列中間插入無編碼作用的鹼基序列,這類基因被稱為間隔或斷裂基因。這一發現於1977年被英國的查弗里斯和荷蘭的弗蘭威爾在研究兔β-球蛋白結構時所證實。1978年,生化學家沃特·吉爾伯特提出基因是一個轉錄單位的設想,他認為基因是一個DNA序列的嵌合體,同時包含兩個區段:一個區段將被表達並存在於成熟的mRNA中,稱為「外顯子」;一個區段由雖然也同時被表達,但將在成熟mRNA中被刪除,稱為「內含子」。近年來的研究發現,原核生物的基因序列一般是連續的,在一個基因的內部幾乎不含「內含子」,而真核生物中絕大多數基因都是由不連續DNA序列組成的斷裂基因。斷裂基因的表達過程是:整個基因先由DNA轉錄成一條信息RNA前體(precursor mRNA),其中的內含序列會被一種稱為「剪接體」的RNA/蛋白質複合物所切除,兩端再相互連接成一條連續的核酸順序,以形成成熟的mRNA。DNA分子斷裂基因的存在為基因功能的展現賦予了更大的潛力。
★重疊基因:
長期以來,人們一直認為在同一段DNA序列內是不可能存在重疊的讀碼結構的。但是,1977年,維納(Weiner)在研究Q0病毒的基因結構時,首先發現了基因的重疊現象。1978年,費爾(Feir)和弗雷德里克·桑格在研究分析φX174噬菌體的核苷酸序列時,也發現由5375個核苷酸組成的單鏈DNA所包含的10個基因中有幾個基因具有不同程度的重疊,但是這些重疊的基因具有不同的讀碼框架。以後在噬菌體G4、MS2和SV40中都發現了重疊基因。基因的重疊性使有限的DNA序列包含了更多的遺傳信息,是生物對它的遺傳物質經濟而合理的利用。
★假基因(偽基因):
1977年,G·Jacp在對非洲爪贍5SrRNA基因簇的研究後提出了假基因的概念,這是一種核苷酸序列同其相應的正常功能基因基本相同,但卻不能合成出功能蛋白質的失活基因。假基因的發現是真核生物應用重組DNA技術和序列分析的結果。現已在大多數真核生物中發現了假基因,如Hb的假基因、干擾素、組蛋白、α球蛋白和β球蛋白、肌動蛋白及人的rRNA和tRNA基因均含有假基因。由於假基因不工作或無效工作,故有人認為假基因,相當人的痕跡器官,或作為後補基因。
★移動基因:
1950年,美國遺傳學家麥克林托卡在玉米染色體組中首先發現移動基因。她發現玉米染色體上有一種稱為Ds的控制基因會改變位置,同時引起染色體斷裂,使其離開或插入部位鄰近的基因失活或恢復恬性,從而導致玉米籽粒性狀改變。這一研究當時並沒有引起重視。20世紀60年代未,英國生物化學家夏皮羅和前西德生物化學家西特爾分別在細菌中發現一類稱為插入順序的可移動位置的遺傳因子,20世紀70年代早期又發現細菌質體的某些抗藥性可移動的基因,到20世紀80年代已發現這類基因至少有20種。20世紀90年代之前,科學家終於用實驗證明了麥克林托卡的觀點,移動基因不僅能在個體的染色體組內移動,並能在個體間甚至種間移動。現已了解到真核細胞中普遍存在移動基因。基因移動性的發現不僅打破了遺傳的DNA恆定論,而且對於認識腫瘤基因的形成和表達,以及生物演化中信息量的擴大等研究工作也將提供新的啟示和線索。
根據操縱子學說,並不是所有的基因都能為肽鏈進行編碼。於是便把能為多肽鏈編碼的基因稱為結構基因,包括編碼結構蛋白和酶蛋白的基因,也包括編碼阻遏蛋白或激活蛋白的調節基因。有些基因只能轉錄而不能轉譯,如tRNA基因和rRNA基因。還有些DNA區段,其本身並不進行轉錄,但對其鄰近的結構基因的轉錄起控制作用,被稱為啟動基因和操縱基因。啟動基因、操縱基因與其控制下的一系列結構基因組成一個功能單位叫做操縱子(operon)。就其功能而言,調節基因、操縱基因和啟動基因都屬於調控基因。這些基因的發現,大大拓寬了人們對基因功能及相互關係的認識。
★斷裂基因:
20世紀70年代中期,法國生物化學家查姆幫(Chamobon)和波蓋特(Berget)在研究雞卵清蛋白基因的表達中發現,細胞內的結構基因並非全部由編碼序列組成,而是在編碼序列中間插入無編碼作用的鹼基序列,這類基因被稱為間隔或斷裂基因。這一發現於1977年被英國的查弗里斯和荷蘭的弗蘭威爾在研究兔β-球蛋白結構時所證實。1978年,生化學家沃特·吉爾伯特提出基因是一個轉錄單位的設想,他認為基因是一個DNA序列的嵌合體,同時包含兩個區段:一個區段將被表達並存在於成熟的mRNA中,稱為「外顯子」;一個區段由雖然也同時被表達,但將在成熟mRNA中被刪除,稱為「內含子」。近年來的研究發現,原核生物的基因序列一般是連續的,在一個基因的內部幾乎不含「內含子」,而真核生物中絕大多數基因都是由不連續DNA序列組成的斷裂基因。斷裂基因的表達過程是:整個基因先由DNA轉錄成一條信息RNA前體(precursor mRNA),其中的內含序列會被一種稱為「剪接體」的RNA/蛋白質複合物所切除,兩端再相互連接成一條連續的核酸順序,以形成成熟的mRNA。DNA分子斷裂基因的存在為基因功能的展現賦予了更大的潛力。
★重疊基因:
長期以來,人們一直認為在同一段DNA序列內是不可能存在重疊的讀碼結構的。但是,1977年,維納(Weiner)在研究Q0病毒的基因結構時,首先發現了基因的重疊現象。1978年,費爾(Feir)和弗雷德里克·桑格在研究分析φX174噬菌體的核苷酸序列時,也發現由5375個核苷酸組成的單鏈DNA所包含的10個基因中有幾個基因具有不同程度的重疊,但是這些重疊的基因具有不同的讀碼框架。以後在噬菌體G4、MS2和SV40中都發現了重疊基因。基因的重疊性使有限的DNA序列包含了更多的遺傳信息,是生物對它的遺傳物質經濟而合理的利用。
★假基因(偽基因):
1977年,G·Jacp在對非洲爪贍5SrRNA基因簇的研究後提出了假基因的概念,這是一種核苷酸序列同其相應的正常功能基因基本相同,但卻不能合成出功能蛋白質的失活基因。假基因的發現是真核生物應用重組DNA技術和序列分析的結果。現已在大多數真核生物中發現了假基因,如Hb的假基因、干擾素、組蛋白、α球蛋白和β球蛋白、肌動蛋白及人的rRNA和tRNA基因均含有假基因。由於假基因不工作或無效工作,故有人認為假基因,相當人的痕跡器官,或作為後補基因。
★移動基因:
1950年,美國遺傳學家麥克林托卡在玉米染色體組中首先發現移動基因。她發現玉米染色體上有一種稱為Ds的控制基因會改變位置,同時引起染色體斷裂,使其離開或插入部位鄰近的基因失活或恢復恬性,從而導致玉米籽粒性狀改變。這一研究當時並沒有引起重視。20世紀60年代未,英國生物化學家夏皮羅和前西德生物化學家西特爾分別在細菌中發現一類稱為插入順序的可移動位置的遺傳因子,20世紀70年代早期又發現細菌質體的某些抗藥性可移動的基因,到20世紀80年代已發現這類基因至少有20種。20世紀90年代之前,科學家終於用實驗證明了麥克林托卡的觀點,移動基因不僅能在個體的染色體組內移動,並能在個體間甚至種間移動。現已了解到真核細胞中普遍存在移動基因。基因移動性的發現不僅打破了遺傳的DNA恆定論,而且對於認識腫瘤基因的形成和表達,以及生物演化中信息量的擴大等研究工作也將提供新的啟示和線索。
★基因的突變★
基因突變和許多疾病的發生有關,如與腫瘤發生有關的癌基因和腫瘤抑制基因。
★從染色體的角度來分類
1.缺失
2.重複
3.倒位
4.易位
★以功能分類
1.失去功能的突變
2.次形態突變
3.超形態突變
4.獲得功能的突變
★以突變原理分類
1.點突變
2.沉默突變
3.錯義突變
4.移碼突變
5.無義突變
★從染色體的角度來分類
1.缺失
2.重複
3.倒位
4.易位
★以功能分類
1.失去功能的突變
2.次形態突變
3.超形態突變
4.獲得功能的突變
★以突變原理分類
1.點突變
2.沉默突變
3.錯義突變
4.移碼突變
5.無義突變
★基因與去氧核甘酸的關係★
(1)基因的基本組成單位是去氧核苷酸。
(2)基因中去氧核苷酸的排列順序稱為遺傳信息。
(3)基因中去氧核苷酸的排列順序的多樣性決定了基因的多樣性。
(2)基因中去氧核苷酸的排列順序稱為遺傳信息。
(3)基因中去氧核苷酸的排列順序的多樣性決定了基因的多樣性。
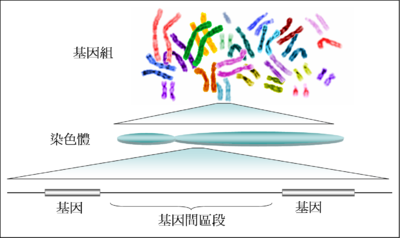
★基因與DNA的關係★
(1)基因是有遺傳效應的DNA片段,每個DNA分子上有許多個基因。一個DNA分子上的鹼基總數大於 該DNA分子上所有基因上的鹼基數之和。
(2)基因具有遺傳效應是指其能控制生物的性狀。基因是控制生物性狀的結構和功能的基本單位,特 定的基因控制特定的性狀。
(2)基因具有遺傳效應是指其能控制生物的性狀。基因是控制生物性狀的結構和功能的基本單位,特 定的基因控制特定的性狀。
★基因與染色體的關係★
(1)基因在染色體上呈線性排列。
(2)染色體是基因的主要載體,但不是唯一載體,如粒線體,葉綠體中也有少量的DNA,也是基因的 載體。
(2)染色體是基因的主要載體,但不是唯一載體,如粒線體,葉綠體中也有少量的DNA,也是基因的 載體。
